Jenouvrier S. (1)*, Brooks C. (2), DuVivier A. (3), and Trathan P.N. (4).
(1) Biology Department, Woods Hole Oceanographic Institution, Woods Hole, MA, USA. *[email protected]
(2) Department of Environmental Studies, University of Colorado Boulder, Boulder, CO, USA.
(3) National Center for Atmospheric Research, Boulder, Colorado, USA.
(4) School of Ocean and Earth Science, University of Southampton, National Oceanography Centre Southampton, European Way, Southampton, United Kingdom.
Translated by: Maximiliano Anzibar Fialho and Luis Cardona.
- El ciclo vital del pingüino emperador está ligado al hielo marino antártico, y las poblaciones sólo prosperan dentro de un rango óptimo de abundancia de hielo marino.
- El tamaño de las colonias individuales de pingüinos emperador varía notablemente, con algunas mostrando un descenso drástico y otras aumentando de tamaño.
- Según un modelo metapoblacional que tiene en cuenta la abundancia del hielo marino y la disminución del mismo proyectada por los modelos Earth System bajo diferentes escenarios de emisiones, casi todas las colonias de pingüinos emperador se habrán extinguido en el año 2100.
- Aunque pequeñas, se siguen descubriendo nuevas colonias de pingüinos emperador gracias a los avances de la teledetección por satélite de ultra alta resolución que amplían la escala espacial del monitoreo.
- No obstante, el índice de abundancia de la población mundial de pingüinos emperador adultos sugiere un descenso del 9,6% entre 2009 y 2018.
- Este declive de la población es más rápido de lo previsto y aún no se comprende del todo, enfatizando la necesidad de realizar esfuerzos urgentes para llevar a cabo un monitoreo más exhaustivo mediante la colaboración internacional.
Ciclo de vida
El pingüino emperador (Aptenodytes forsteri) tiene un ciclo vital especializado que está íntimamente ligado al hielo marino antártico, permitiéndole sobrevivir durante seis meses de oscuridad y temperaturas gélidas en el lugar más frío y seco de la Tierra (Ainley et al. 2010, ver el vídeo El ciclo vital de los pingüinos emperador está ligado al hielo marino). Los ejemplares de esta especie dependen del hielo marino para reproducirse, descansar, mudar las plumas, alimentarse y buscar refugio de los depredadores. Su relación con el hielo marino depende de un delicado equilibrio; una cantidad insuficiente de hielo marino afecta a las redes tróficas marinas y reduce la disponibilidad de alimentos, además de impedir que los pingüinos encuentren un hábitat estable para reproducirse (en particular, el hielo fijo, que es el hielo marino que está pegado a la costa o a plataformas de hielo, Massom et al. 2009); por el contrario, una cantidad excesiva de hielo marino aumenta la distancia de viaje entre las zonas de nidificación y de alimentación, lo que limita su capacidad para alimentar adecuadamente a sus polluelos (véase Hielo marino antártico n.º 1: papel físico y función).
El pingüino emperador es la única especie de sangre caliente que se reproduce en la Antártida en pleno invierno. Los pingüinos llegan a sus lugares de cría en marzo o abril, tras el inicio de la formación del hielo marino durante el otoño austral (Prevost 1961). A mediados de mayo, las hembras ponen un solo huevo y luego realizan un largo viaje para buscar alimento en aguas abiertas. Losl machos incuban un único huevo durante 65 a 75 días a lo largo del invierno antártico, mientras las hembras están fuera. Durante este periodo, los machos debe sobrevivir sin comer. Cuando las hembras regresan al final del invierno para alimentar a los polluelos recién nacidos, los machos han perdido casi la mitad de su peso corporal, tras ayunar durante cuatro meses.
Para sobrevivir al frío y a las frecuentes y violentas tormentas y ventiscas del invierno, los pingüinos macho se amontonan para calentarse. Cientos de pingüinos se agrupan, cambiando continuamente de posición dentro del grupo. Así, cada individuo intenta continuamente evitar la parte externa más fría del grupo, beneficiándose de las partes centrales, más cálidas. Este movimiento coordinado contribuye a mantener una temperatura ambiente en el interior del grupo superior a 0 °C, incluso con una temperatura exterior media de -17 °C; el interior protegido del grupo puede alcanzar los 37,5 °C (Gilbert et al. 2006).
Las hembras regresan en agosto, justo después de la eclosión de los huevos. Ambos progenitores se turnan entonces, alternando la búsqueda de comida y el aprovisionamiento del polluelo, además de mantenerlo caliente. En septiembre, ambos adultos deben dejar simultáneamente al polluelo para poder buscar comida y satisfacer su creciente demanda. En diciembre, adultos y polluelos abandonan la colonia y regresan al océano. Los adultos deben entonces alimentarse antes de la muda, durante la cual reemplazaran todas sus plumas. Durante la muda, los adultos no se alimentan y necesitan una plataforma estable de hielo marino, ya que su plumaje ya no es adecuado para aislarlos del agua fría.
Para alimentarse, los pingüinos emperador pueden sumergirse durante casi 30 minutos con el aire una sola inspiración, por lo que sus inmersiones son más largas y profundas (hasta un máximo de 546 m, aunque la mayoría son menos profundas (Wienecke et al. 2007)) que las de cualquier otra especie de ave. Los pingüinos emperador se alimentan principalmente de diablillo antártico, krill antártico, calamar flecha y neocalamar antártico, y las condiciones del hielo marino influyen mucho en la disponibilidad de krill y diablillo plateado (véase la revisión de Trathan et al. 2020).
Comprender el presente: La ecología del pingüino emperador y su relación con las condiciones del hielo marino
Los conocimientos sobre las tendencias de la población de pingüinos emperador han mejorado gracias tanto a los estudios a largo plazo (Barbraud y Weimerskirch 2001, Wienecke 2010), así como a los avances en teledetección por satélite de alta resolución que permiten detectar sus colonias (Fretwell y Trathan, 2009).
El ciclo vital del pingüino emperador está ligado al hielo marino: implicaciones para las fluctuaciones en el tamaño de las colonias

Figura 1. Mapa que ilustra la ubicación de las colonias existentes y el tamaño medio de sus poblaciones entre 2009 y 2018. Los números de las colonias se refieren a los nombres de los sitios enumerados en la Tabla 1. Las colonias resaltadas en rojo representan aquellas con estimaciones disponibles del tamaño de la población según La Rue et al. 2024. Las colonias marcadas en verde son localizaciones adicionales mencionadas en la bibliografía, incluyendo las referencias de Fretwell y Trathan 2020 y Fretwell 2024. El recuadro del gráfico en el continente antártico muestra el índice de abundancia global de pingüinos emperador, calculado anualmente (La Rue et al 2024). Este cálculo implica combinar el área de "píxeles con pingüinos" de las imágenes de Muy Alta Resolución (VHR) enmarcado en un modelado bayesiano durante el periodo de estudio de 2009 a 2018. Los índices de abundancia de pingüinos emperador pasaron de aproximadamente 252.000 aves presentes durante la primavera de 2009 a aproximadamente 226.000 aves en 2018. La banda roja del gráfico representa el intervalo de credibilidad del 95% de colas iguales para el índice anual, mientras que la línea roja central muestra la mediana de la distribución posterior. La línea negra discontinua muestra la tendencia lineal.
| Site # | Site-Name | Latitude | Longitude | Count | Color on map |
| 1 | Snow Hill Island | -64.5238 | -57.4445 | 3324 | Red |
| 2 | Jason Peninsula | -66.0996 | -60.6737 | 1788 | Red |
| 3 | Gipps Ice Rise | -68.68 | -60.86 | 0 | Green |
| 4 | Dolleman | -70.6106 | -60.4207 | 3771 | Red |
| 5 | Cape Darlington | -71.8872 | -60.1338 | 0 | Green |
| 6 | Smith | -74.3693 | -60.8273 | 3198 | Red |
| 7 | Gould | -77.7099 | -47.6562 | 4424 | Red |
| 8 | Luitpold | -77.271 | -33.552 | 4614 | Red |
| 9 | Halley | -75.54 | -27.43 | 12337 | Red |
| 10 | Dawson | -76.0143 | -26.6479 | 4307 | Red |
| 11 | Stancomb | -74.1203 | -23.0867 | 6012 | Red |
| 12 | Drescher | -72.8257 | -19.3262 | 7365 | Red |
| 13 | Riiser | -72.1245 | -15.106 | 4970 | Red |
| 14 | Atka | -70.6142 | -8.1317 | 6831 | Red |
| 15 | Sanae | -69.9994 | -1.4128 | 4416 | Red |
| 16 | Astrid | -69.9481 | 8.3176 | 3252 | Red |
| 17 | Lazarev Ice Shelf | -69.38 | 14.64 | 0 | Green |
| 18 | Lazarev | -69.7504 | 15.5493 | 217 | Red |
| 19 | Ragnhild | -69.908 | 27.1548 | 4626 | Red |
| 20 | Gunnerus | -68.7625 | 34.3819 | 2789 | Red |
| 21 | Umbeashi | -68.0459 | 43.0172 | 112 | Red |
| 22 | Casey Bay West | -67.312 | 46.957 | 0 | Green |
| 23 | Amundsen Bay | -66.7826 | 50.5437 | 69 | Red |
| 24 | Kloa Point | -66.6407 | 57.2779 | 1289 | Red |
| 25 | Fold Island | -67.3239 | 59.3159 | 287 | Red |
| 26 | Taylor Glacier | -67.4543 | 60.8778 | 798 | Red |
| 27 | Auster Islands | -67.3966 | 63.9742 | 4035 | Red |
| 28 | Cape Darnley | -67.887 | 69.696 | 2664 | Red |
| 29 | Amanda Bay | -69.2711 | 76.8348 | 4267 | Red |
| 30 | Barrier Bay | -66.55 | 81.818 | 359 | Red |
| 31 | West Ice Shelf | -67.225 | 81.931 | 0 | Green |
| 32 | Karelin Bay | -66.4118 | 85.3836 | 0 | Green |
| 33 | West Ice Shelf East | -66.08 | 86.53 | 0 | Green |
| 34 | Burton Ice Shelf | -66.272 | 89.695 | 0 | Green |
| 35 | Haswell Island | -66.5308 | 93.0079 | 3289 | Red |
| 36 | Shackleton Ice Shelf | -65.0887 | 96.02 | 4083 | Red |
| 37 | Bowman Island | -65.1613 | 103.0674 | 1320 | Red |
| 38 | Peterson Bank | -65.9175 | 110.2355 | 1553 | Red |
| 39 | Cape Poinsett | -65.7817 | 113.2351 | 0 | Green |
| 40 | Sabrina Coast | -66.177 | 121.058 | 0 | Green |
| 41 | Porpoise Bay | -66.3204 | 129.7496 | 0 | Green |
| 42 | Dibble Glacier | -65.9999 | 134.7996 | 9711 | Red |
| 43 | Point Geologie | -66.6744 | 140.0052 | 1578 | Red |
| 44 | Mertz Glacier Break-Off | -67.322 | 145.893 | 0 | Green |
| 45 | Mertz Glacier | -66.892 | 146.62 | 5292 | Red |
| 46 | Ninnis Bank | -66.7229 | 149.677 | 0 | Green |
| 47 | Davis Bay | -69.3484 | 158.4924 | 2212 | Red |
| 48 | Yule Bay | -70.7161 | 166.4777 | 0 | Green |
| 49 | Cape Roget | -71.9882 | 170.5972 | 7983 | Red |
| 50 | Coulman Island | -73.3483 | 169.6242 | 26477 | Red |
| 51 | Cape Washington | -74.6373 | 165.3821 | 14040 | Red |
| 52 | Franklin Island | -76.1873 | 168.4401 | 6348 | Red |
| 53 | Beaufort Island | -76.94 | 167.039 | 1416 | Red |
| 54 | Cape Crozier | -77.4653 | 169.3294 | 1071 | Red |
| 55 | Cape Colbeck, Edward VII Peninsula | -77.14 | -157.7 | 22266 | Red |
| 56 | Rupert Coast | -75.3821 | -143.3076 | 1753 | Red |
| 57 | Cruzen Island | -74.7341 | -140.3266 | 0 | Green |
| 58 | Verleger Point | -74.71 | -136.07 | 0 | Green |
| 59 | Ledda Bay | -74.42 | -130.96 | 616 | Red |
| 60 | Thurston Glacier, Mount Siple | -73.4977 | -125.6204 | 3403 | Red |
| 61 | Cape Gates | -73.6609 | -122.697 | 0 | Green |
| 62 | Bear Peninsula | -74.3922 | -110.1917 | 3684 | Red |
| 63 | Brownson Islands | -74.14 | -103.48 | 3872 | Red |
| 64 | Noville Peninsula | -71.7693 | -98.4467 | 3046 | Red |
| 65 | Pfrogner Point | -72.5687 | -89.9058 | 0 | Green |
| 66 | Bryan Coast | -73.2491 | -85.3477 | 1036 | Red |
| 67 | Smyley | -72.3017 | -78.8195 | 4314 | Red |
| 68 | Verdi Inlet | -71.5559 | -74.7603 | 0 | Green |
| 69 | Rothschild Island | -69.5209 | -72.2293 | 797 | Red |
| 70 | Emperor Island | -67.8647 | -68.71 | 0 | Green |
Tabla 1. Descripción de la ubicación de las colonias de pingüinos emperador y su índice de abundancia poblacional promediado según La Rue et al. 2024. Los números se refieren a la Figura 1.
Tras el inicio del monitoreo rutinario mediante microondas pasivas por satélite en 1979, la extensión del hielo marino en toda la Antártida mostró un ligero aumento neto hasta 2016, aunque las tendencias variaron notablemente según la región y la estación (Parkinson 2019). Sin embargo, en los últimos años se han producido descensos drásticos en la extensión del hielo marino hasta alcanzarse niveles mínimos históricos (Turner et al. 2022, véase Hielo marino antártico n.º 3: Tendencias y proyecciones futuras). Los factores atmosféricos y oceánicos cambiantes, incluidos los vientos más rápidos y el enfriamiento y calentamiento del océano (Hobbs et al. 2016, Purich y Doddridge 2023), han influido en estas tendencias del hielo marino.
Tanto la mortalidad como el éxito reproductivo del pingüino emperador dependen en gran medida de las condiciones del hielo marino y existe un rango óptimo de hielo marino en el que las poblaciones prosperan (Jenouvrier et al. 2012). Por ejemplo, a finales de la década de 1970, la población de la colonia de Pointe Géologie (#43 Figura 1) se redujo a la mitad, debido a un incremento en la la mortalidad de los machos durante varios años consecutivos debido a la limitada extensión del hielo marino (Barbraud y Weimerskirch 2001, Jenouvrier et al. 2005). La baja extensión del hielo marino reduce la abundancia de especies de presa clave, siend los machos más vulnerables que las hembras debido a su largo ayuno invernal. Además, la población de la colonia de Pointe Géologie también ha experimentado fracasos de cría masivos en 1982, 1990, 1992, 1994, 1995, 2013 y 2014 (Jenouvrier et al. 2009, Barbraud et al. 2015) cuando el aumento de la extensión del hielo marino incrementó la distancia entre las aguas abiertas más cercanas y la colonia durante el periodo de cría de los polluelos (Massom et al. 2009; Labrousse et al. 2021). Estos impactos en la población son importantes, ya que el pingüino emperador tiene un periodo generacional de 16 años (Jenouvrier et al. 2014), por lo que las alteraciones en la reproducción y el éxito reproductivo tardan mucho tiempo en estabilizarse.
Los cambios interanuales en el tamaño de la población de una colonia determinada varían notablemente, con algunas colonias mostrando descensos drásticos, mientras que otras colonias cercanas registran aumentos notables. No todas esas variaciones pueden comprenderse fácilmente (por ejemplo, Barber-Mayer et al. 2008), pero algunos cambios importantes en la población pueden vincularse a perturbaciones físicas importantes (por ejemplo, Kooyman et al. 2007). Así, en la segunda colonia de pingüinos emperador más grande del mundo (Halley Bay, #9 Figura 1), la población reproductora disminuyó a prácticamente cero en 2015, con consecuencias para la cercana colonia de Dawson-Lambton (#10 Figura 1), a solo 55 km al sur, que experimentó un aumento de más de diez veces en el número de pingüinos (Fretwell y Trathan 2019). Halley Bay sufrió tres años de fracaso casi total en la cría debido a un cambio en las condiciones atmosféricas y, por tanto, en el hielo marino. Es casi seguro que esas malas condiciones provocaron la reubicación de los pingüinos, que en su mayoría se trasladaron a Dawson-Lambton en los años siguientes. En otros lugares, también se cree que los pingüinos cambian de lugar de cría en algunos años (Kooyman y Ponganis, 2017), mientras que algunas colonias desaparecen en ciertos años y reaparecen en los años siguientes (Fretwell y Trathan 2021), lo que pone de relieve la complejidad de los factores que impulsan las tendencias de la población y la idoneidad del hábitat para el pingüino emperador.
Distribución panantártica del pingüino emperador y dinámica de la población mundial
Actualmente se han identificado setenta colonias de pingüinos emperador alrededor de la Antártida (Fretwell y Trathan 2019, Fretwell y Trathan 2021, Fretwell 2024, Figura 1 y Tabla 1). Utilizando imágenes satelitales circumpolares de alta resolución, métricas relacionadas con la extensión del hielo fijo y otros factores geográficos y biológicos, Labrousse et al. (2023) revelaron que no existen diferencias discernibles entre los hábitats de hielo fijo ocupados por colonias de pingüinos y aquellos que carecen de ellas. No obstante, las polinias persistentes (Massom et al. 1998), las grietas en el hielo marino, las fallas y las polinias efímeras de corta duración (Labrousse et al. 2019), pueden ser características clave del hábitat que proporcionen acceso a potenciales zonas de alimentación adecuadas para el pingüino emperador.
Recientemente, el descubrimiento de nuevas colonias ha aumentado la estimación de la población global, al tiempo que se ha ampliado la escala geográfica de seguimiento, incluyendo ahora zonas alejadas de los márgenes costeros (Fretwell y Trathan 2021, Fretwell 2024). Además, LaRue et al. (2024) han combinado la teledetección con encuestas de validación y la modelización bayesiana para evaluar el tamaño de la población y la trayectoria de los pingüinos emperador adultos en toda el área de distribución de la población. El nuevo análisis estima un índice global de abundancia de 252.000 adultos reproductores, con una disminución media estimada de la población del 9,6% (intervalo de credibilidad del 95%: -26,4% a +9,4%) entre 2009 y 2018 (LaRue et al. 2024). Existen importantes fluctuaciones interanuales en torno a esta tendencia, mostrando un descenso inicial, seguido posteriormente por un aparente aumento de la población hacia la última parte de la serie temporal; sin embargo, la modelización sugiere una probabilidad del 81% de un descenso general. Además, la posibilidad de emigración hacia y desde colonias fuera de las 50 colonias examinadas por el estudio de LaRue et al. (2024) podría contribuir a la disminución observada durante el período 2009-2018. Esta y otras explicaciones no pueden descartarse por completo. Los mecanismos ecológicos que subyacen a los cambios observados no se comprenden del todo, lo que subraya la necesidad de realizar esfuerzos urgentes para realizar un monitoreo más amplio mediante la colaboración internacional.
Esta nueva modelización de la población tiene en cuenta las distintas tendencias a nivel de colonia, la desaparición y reaparición ocasional de colonias y la variación diaria de la abundancia durante el periodo de estudio de primavera. La modelización también tiene en cuenta las imperfecciones en los recuentos de las prospecciones aéreas y las imprecisas observaciones por satélite de las zonas ocupadas, considerando los posibles cambios en los recuentos previstos a lo largo del periodo de prospección. Es importante señalar que el nuevo índice sólo incluye una proporción de la población presente a finales de primavera (los adultos reproductores), por lo que proporciona una estimación conservadora del número total de pingüinos emperador adultos, ya que no incluye a los reproductores fracasados ni a las aves jóvenes que aún no han comenzado a reproducirse.
Este índice revela una disminución media anual de la población mundial del -1,3% entre 2009 y 2018 (IC del 95% = -3,3% a +1,0%) y sugiere que probablementese hayan producido disminuciones en cuatro de las ocho regiones de hielo fijo, independientemente de las condiciones del hábitat. La mayoría de los lugares de cría del pingüino emperador se encuentran en el hielo fijo y si este se rompe demasiado pronto en la temporada, antes de que los polluelos se hayan desarrollado completamente y dispongan ya de plumas impermeables, estos pueden ahogarse en el agua, lo que lleva a fracasos de cría generalizados (Fretwell y Trathan 2019; Fretwell et al. 2023). Las mediciones precisas de la extensión del hielo fijo siguen siendo un reto. La información sobre el cambio a gran escala y la variabilidad en la cobertura de hielo fijo circumantártico se deriva del análisis de series temporales de imágenes de satélite (Fraser et al. 2021). Estos y otros estudios muestran que la extensión, persistencia y estacionalidad del hábitat de hielo fijo están influidas por la presencia/ausencia de un «colchón» protector de hielo compacto (Massom et al. 2009; Fraser et al. 2021). A largo plazo, es probable que el hielo marino a gran escala determine la extensión del hielo fijo como plataforma de reproducción (Fraser et al. 2021).
Anticipar el futuro: Previsión de la dinámica del pingüino emperador bajo el cambio climático
En futuros escenarios de cambio climático con temperaturas globales más cálidas (IPCC, 2023- Lee at al. 2023), el hielo marino antártico se formará más tarde y se romperá antes que en la actualidad. A pesar de las incertidumbres relativas a la magnitud de los cambios futuros en el hielo marino, los modelos del sistema terrestre acoplados (ESMs) proyectan sistemáticamente la pérdida de hielo marino antártico en todas las estaciones (Fox-Kemper et al. 2021).
Las proyecciones de población de pingüino emperador se determinan utilizando modelos demográficos de última generación que incluyen las complejas relaciones entre reproducción, mortalidad y hielo marino a lo largo de todo el ciclo vital de los pingüinos emperador, y están estrechamente vinculadas con las proyecciones de hielo marino derivadas de los ESMs (Jenouvrier et al. 2012, 2013, 2014, 2017, 2020, 2021). Actualmente, estos modelos demográficos no contemplan el impacto del hielo fijo en la reproducción, ya que las proyecciones de hielo fijo no están disponibles para ningún ESMs; sin embargo, la estabilidad del hielo fijo está fuertemente influenciada por el hielo marino adyacente (Fraser et al. 2021), por lo que a medida que el hielo marino disminuye, es probable que el hielo fijo también lo haga. Es prácticamente seguro que la disminución prevista del hielo marino provoque una reducción de la población mundial de pingüinos emperador (Jenouvrier et al. 2014, 2021). De hecho, es probable que todas las colonias de pingüinos emperador disminuyan a finales de siglo, independientemente del escenario de emisiones de gases de efecto invernadero, con o sin mitigación de emisiones.
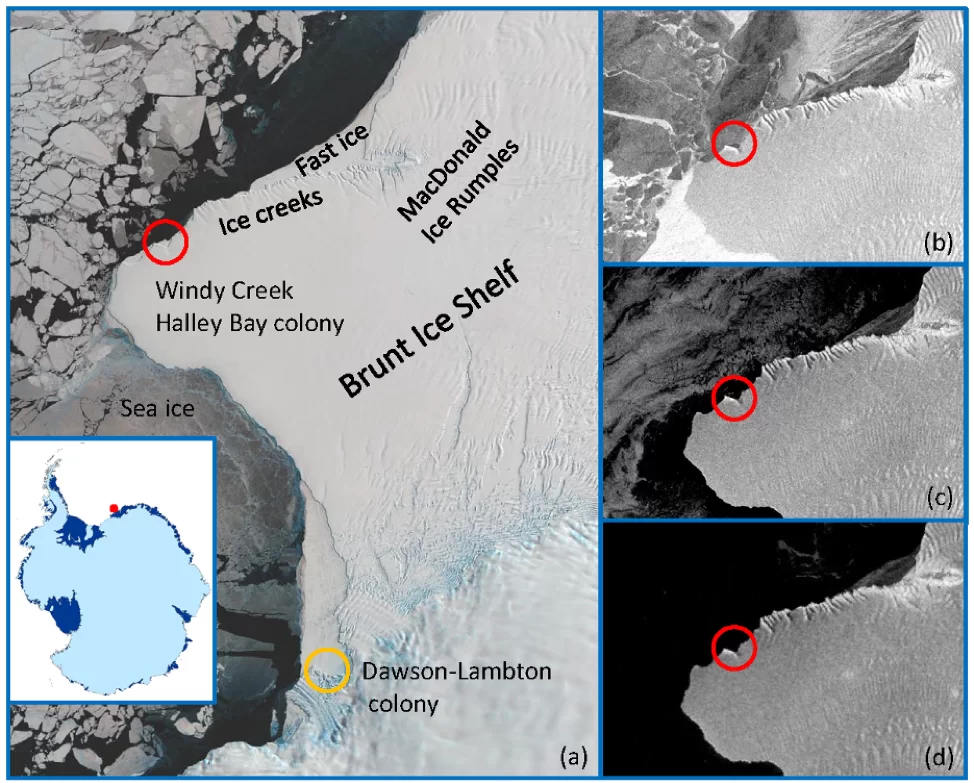
Figura 2. (a) Imagen Landsat8 de octubre de 2016 que muestra la ubicación de la colonia de Halley Bay en Windy Creek, junto con la ubicación de la colonia Dawson-Lambton en el glaciar del mismo nombre. Área alrededor de Windy Creek el (b) 3 de noviembre de 2016; (c) 17 de noviembre de 2016; y (d) 1 de diciembre de 2016 en imágenes obtenidas del satélite Copernicus Sentinel 1 de la Unión Europea, utilizando el EO Browser. En 2016, la reproducción fracasó en Halley Bay debido a la pérdida de hielo marino, tal como se muestra en los paneles b-d; se cree que muchas aves se trasladaron a Dawson-Lambton en años posteriores. Obsérvese que la plataforma de hielo Brunt experimentó un importante evento de desprendimiento en enero de 2023, y aún no se sabe con certeza si los pingüinos emperador han comenzado a regresar (Comunicado de prensa British Antarctic Survey, 23 de enero de 2023).
Además de analizar los resultados medios a largo plazo, los científicos también han tenido en cuenta el efecto de fenómenos medioambientales extremos en sus modelos de población (Jenouvrier et al. 2021). Por ejemplo, las imágenes por satélite mostraron que en 2016 se produjo una formación tardía y una ruptura temprana del hielo marino sobre el que se encontraba la colonia de Halley Bay (Figura 2), y probablemente murieron más de 10.000 polluelos (Fretwell y Trathan 2019). Otros fenómenos extremos, como el desprendimiento de glaciares, lenguas de hielo y plataformas de hielo, tienen el potencial de socavar la estabilidad del hielo fijo, haciendo necesaria la reubicación de las colonias de pingüinos (Fretwell et al. 2014). El desprendimiento de las plataformas de hielo, en particular, puede dar lugar a la creación de grandes icebergs tabulares que pueden impedir el acceso de los pingüinos a las zonas de alimentación. Esta interrupción tiene el potencial de afectar al éxito reproductivo, al aumentar las distancias a las zonas de alimentación y causar daños a las plataformas de cría situadas sobre el hielo fijo (Kooyman et al. 2017).
Además de revelar los efectos de fenómenos medioambientales extremos, las imágenes por satélite también han ayudado a desentrañar otros efectos que influyen en el tamaño de las colonias. Las imágenes por satélite muestran que los movimientos de los pingüinos emperador entre colonias pueden hacer que éstas crezcan o disminuyan. Con este conocimiento, los científicos desarrollaron modelos para incorporar comportamientos de movimiento y evaluar cómo podrían cambiar las poblaciones a medida que los pingüinos se desplazan entre colonias. Estos modelos suponen que los pingüinos abandonarán un hábitat de baja calidad para buscar, a corta y larga distancia, un hábitat de mayor calidad que maximice sus posibilidades de reproducirse con éxito. Sin embargo, ni siquiera considerando las actividades de dispersión y la formación de nuevas colonias, se invertirá el declive poblacional previsto.
Combinando los efectos de la dispersión y los fenómenos extremos en los modelos demográficos del pingüino emperador, Jenouvrier et al. (2021) proyectaron que el 98% de las colonias se extinguirán en 2100 en un escenario de calentamiento climático elevado (escenario «de referencia» RCP8.5, en el que las emisiones de gases de efecto invernadero se mantienen sin cambios durante todo el siglo XXI), y que la población mundial disminuirá en un 99% en comparación con su tamaño histórico (Figura 3). En un escenario que incluya el cumplimiento del Acuerdo de París, y se limite el aumento medio de la temperatura global a no más de 2 °C, con esfuerzos adicionales para restringir el aumento a solo 1,5 °C, el descenso previsto del número de colonias persiste, pero en mucha menor medida. Bajo este escenario, se anticipa que el 61% de las colonias estarán en riesgo de extinción para 2100, desacelerando así, pero no evitando, el declive global de la población (Jenouvrier et al. 2019, Tabla S2 en Jenouvrier et al. 2021 , Figura 3).
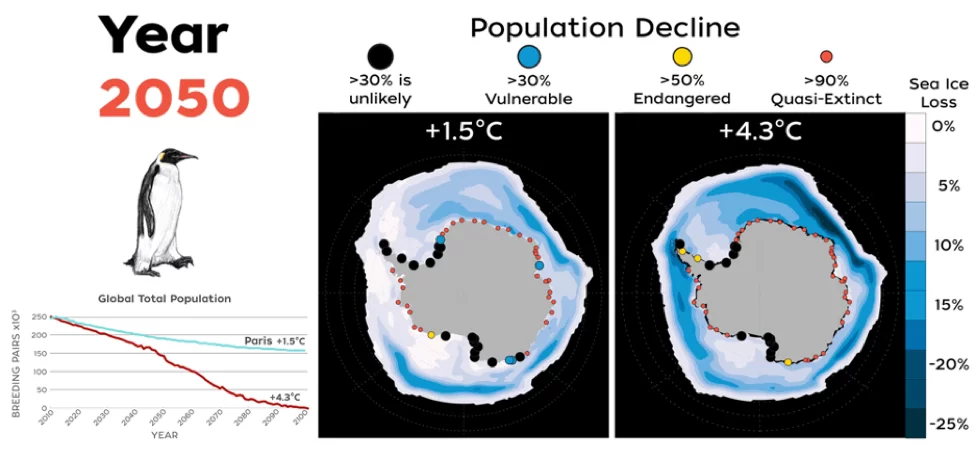
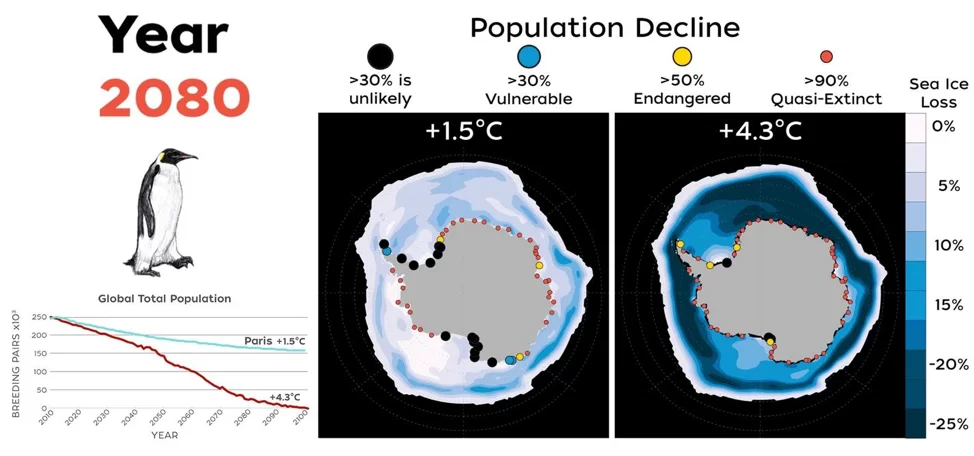
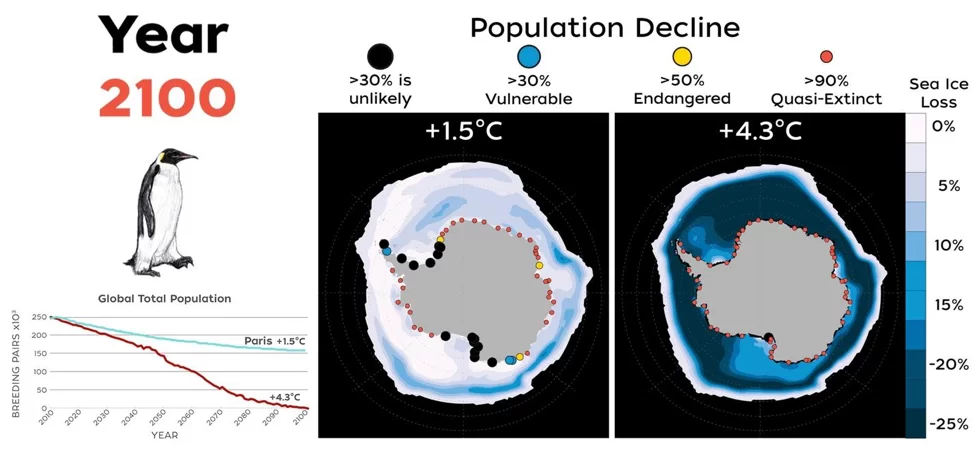
Figura 3. Mapas de colonias de pingüinos emperador y cambios en el hielo marino que contrastan el futuro para un mundo sin o con mitigación climática y el impacto del cambio climático en la población global de pingüinos emperador (actualizado de Jenouvrier et al. 2021). En los mapas, la pérdida de hielo marino se representa desde una pérdida severa en 2100 en relación con la actualidad, en azul oscuro, hasta cambios mucho menores en el hielo marino, en blanco. El color y el tamaño de los puntos muestran el declive de la población en las fechas previstas de 2050, 2080 y 2100. Panel derecho (sin mitigación climática): La mayoría de las colonias habrán desaparecido en un escenario en el que las emisiones de gases de efecto invernadero sigan su curso actual (RCP 8,5, Riahi et al 2011). El 98% de las colonias se habrán prácticamente extinguido a finales de siglo (puntos rojos). Cabe resaltar que las pocas colonias que no hayan desaparecido, mostradas aquí en negro, estarán en declive. Panel izquierdo (con mitigación climática): Si se toman medidas para reducir las emisiones de gases de efecto invernadero y se cumplen los objetivos del Acuerdo de París (Sanderson et al. 2018), en 2100 seguirán existiendo refugios viables para el pingüino emperador en la Antártida. El modelo sigue proyectando descensos, pero las tendencias serán mucho menos graves, de modo que se extinguirán menos colonias. Tendencias de la población mundial: número total de parejas reproductoras de pingüinos emperador de 2009 a 2100 proyectado para varios escenarios climáticos (rojo: las emisiones de gases de efecto invernadero mantienen sus tendencias actuales; azul: los países toman medidas para reducir sus emisiones de gases de efecto invernadero con el fin de alcanzar los objetivos del Acuerdo de París).
Amenazas del cambio climático y retos de la conservación
El Rastreador de la Acción Climática (CAT, https://climateactiontracker.org/) desempeña un papel crucial en la cuantificación y evaluación de los esfuerzos de mitigación del cambio climático, analizando objetivos, políticas y acciones tanto a nivel nacional como mundial. Según las evaluaciones del CAT, las políticas actuales son insuficientes para limitar adecuadamente el calentamiento global. La estimación combinada sugiere ahora un calentamiento medio de 2,7°C (rango plausible entre 2,5°C y 2,9°C) para 2100, con temperaturas que seguirán aumentando más allá de ese punto. Este aumento de la temperatura mundial supone una amenaza importante para los pingüinos emperador. En tales escenarios, la población mundial se habrá reducido a más de la mitad en 2050 (Jenouvrier et al. 2021).
Desplazamiento de especies en la Antártida
Las actividades humanas pueden causar el desplazamiento de especies antárticas nativas a zonas de la Antártida donde no se encuentran de forma natural (transferencia intrarregional) (Hughes et al., 2019; Bergstrom, 2022). La dispersión de especies mediada por el hombre podría perturbar los ecosistemas terrestres y de agua dulce existentes y alterar las distintas regiones biogeográficas que se encuentran dentro de la Antártida (Hughes y Convey, 2010; Terauds y Lee, 2016; Hughes et al., 2019; Cukier et al., 2023). Además, el movimiento humano dentro de la Antártida puede transferir especies exóticas ya existentes a otras zonas antárticas (Hughes et al., 2019). Por ejemplo, las investigaciones de laboratorio han demostrado que un mosquito sin alas (Eretmoptera murphyi), introducido accidentalmente en la isla Signy, en las islas Orcadas del Sur, podría sobrevivir y completar su ciclo vital unos 750 km más al sur, en la península Antártica, y algunas gramíneas podrían sobrevivir más allá de sus distribuciones antárticas actuales (Hughes et al., 2013; Pertierra et al., 2017a).
En conclusión, el pingüino emperador es un ejemplo notable de adaptación a las condiciones extremas del hielo antártico. Sin embargo, modelos avanzados predicen la extinción de la especie para finales de siglo en un escenario de emisiones elevadas, impulsada por la reducción del hábitat de hielo marino causada por el cambio climático. Estos resultados subrayan la urgencia de mejorar la acción climática y las medidas de mitigación para hacer frente a las profundas consecuencias ecológicas del calentamiento global en la Antártida (Lee et al. 2022).