



Phil N. Trathan (1), Heather J. Lynch (2)*, William R. Fraser (3)
* heather.lynch[at]stonybrook.edu
La evidencia sólida destaca cambios importantes en la abundancia y distribución de pingüinos (Adelia, barbijo, papúa, emperador, rey y de penacho anaranjado) que se reproducen en la Península Antártica y en todo el Arco de Scotia, en las Subáreas estadísticas de 48.1 a 48.4 de la FAO. En la Subárea 48.1, los pingüinos Adelia y barbijo han disminuido en la mayor parte de la Península Antártica occidental (WAP) al norte de la bahía Margarita. El pingüino Adelia se encuentra estable o en aumento en la bahía Margarita y al sur, y está estable o en aumento en la Península Antártica oriental. Por el contrario, el pingüino papúa que habita en la WAP (48.1) y en las islas Orcadas (48.2) está aumentando y expandiendo su rango de reproducción hacia el sur; en otros lugares, sus poblaciones son muy variables, aunque no se registran tendencias significativas. En la Subárea 48.3, el pingüino de penacho anaranjado experimentó disminuciones considerables, mientras que el pingüino rey ha aumentado. En la Subárea 48.4, los pingüinos barbijo y Adelia se encuentran estables. Estos hallazgos destacan la considerable heterogeneidad espacial en las tendencias de las especies y la importancia de contar con datos censales actualizados a fin de respaldar el trabajo para la predicción y el seguimiento de las tendencias futuras.
Existen dieciocho especies de pingüinos en el mundo, seis de las cuales se reproducen regularmente en la Península Antártica y en el mar de Scotia (Figura 1): pingüinos Adelia [Pygoscelis adeliae], barbijo [P. antarctica], papúa [P. papua], rey [Aptenodytes patagonicus], emperador [A. forsteri] y de penacho anaranjado [Eudyptes chrysolophus]. Las poblaciones de pingüinos en esta región han cambiado durante el siglo pasado (por ejemplo, 1-2), y se han registrado marcadas disminuciones de muchas colonias de pingüinos de penacho anaranjado, Adelia y barbijos. Por ejemplo, los pingüinos de penacho anaranjado han disminuido en la Subárea 48.3 (3), y, aunque los pingüinos Adelia y barbijos están estables o en aumento en la Subárea 48.1, en la bahía Margarita (2, 4), estos han disminuido en la mayor parte de la Península Antártica occidental (WAP) al norte de la bahía Margarita (2). El pingüino Adelia también se encuentra estable o en aumento en la región del mar de Weddell, así como en la Subárea 48.4 (4). El pingüino papúa tiene un incremento en su abundancia en la WAP en 48.1 y en las islas Orcadas en 48.2, y aumenta su rango de reproducción al sur (2); en otros lugares en la región, las poblaciones de pingüino papúa no parecen mostrar una tendencia significativa (4), aunque las grandes fluctuaciones interanuales en la abundancia de esta especie pueden complicar la evaluación de la tendencia. El pingüino rey ha aumentado en la Subárea 48.3 y parece ampliar su rango hacia el sur de las islas Shetland del Sur en 48.1 (5), y, aunque el pingüino emperador no ha desaparecido de las islas Dion en la WAP (6), continúa reproduciéndose cerca de la isla Cerro Nevado. Estos hallazgos destacan la considerable heterogeneidad espacial en las tendencias poblacionales de pingüinos y la importancia del trabajo comparativo a través de sitios de estudios a largo plazo.
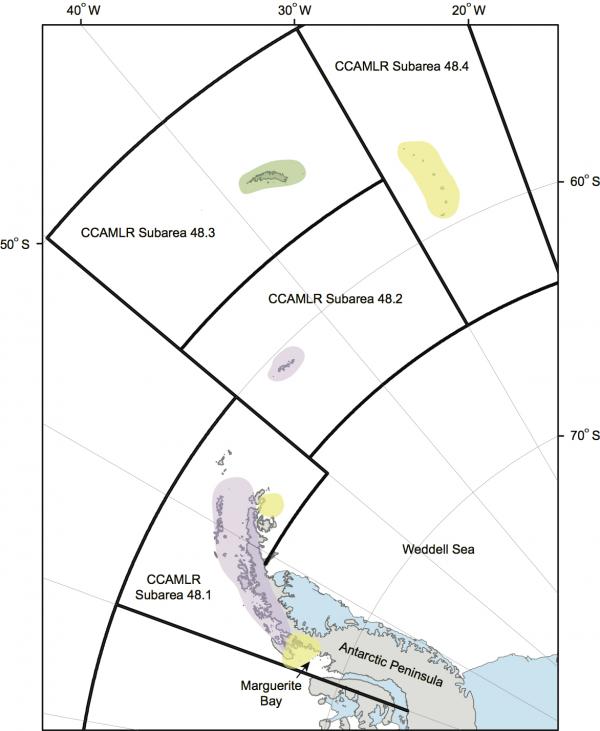
Figura 1. Península Antártica y Arco de Scotia. El área rosa representa esa parte de la Península Antártica occidental donde los pingüinos Adelia y barbijo han disminuido en abundancia y el pingüino papúa ha aumentado en abundancia; las áreas amarillas representan las zonas donde los pingüinos Adelia y barbijo están ya sea estables o aumentan en abundancia (NB: no se encontraron ejemplares de pingüino barbijo en la zona del mar de Weddell ni en la bahía Margarita). El sombreado verde indica la zona donde el pingüino de penacho anaranjado experimentó disminuciones considerables. Las fronteras de las Subáreas estadísticas de la FAO utilizadas por la CCRVMA se muestran en negro.
Conjuntos de datos existentes sobre los pingüinos en la Península Antártica y el Arco de Scotia
Los conjuntos de datos de largo plazo (20 años o más) sobre la abundancia, dieta, demografía y fenología incluyen estudios de las islas Shetland del Sur (por ejemplo, 1,7,8), Puerto Lockroy (9), isla Anvers y punta Biscoe (10), punta Cierva (11), isla Signy (12), isla Avian (13) e isla Bird (3). A través del programa Inventario de Sitios Antárticos (4), que opera a través de la península antártica y las islas Orcadas, se relevaron nidos de pingüinos y polluelos —y otras aves marinas— desde 1994 (4) utilizando embarcaciones turísticas para proporcionar una cobertura geográfica más amplia complementaria a la cobertura proporcionada por otros programas de investigación a largo plazo.
Los datos de población más actualizados y más completos sobre pingüinos antárticos provienen del proyecto MAPPPD (4), una aplicación en la que se almacenan todos los datos publicados sobre poblaciones de pingüinos antárticos, incluida la información histórica del British Antarctic Survey, del programa Inventario de Sitios Antárticos, de la División Antártica Australiana y de otros programas nacionales. Los datos de la MAPPPD también incluyen estimaciones de población hechos a partir de imágenes satelitales de muy alta resolución relativas a pingüinos Adelia (14), recuentos por vehículos aéreos no tripulados, fotografías del terreno y recuentos de embarcaciones en alta mar.
En la Subárea 48.1, los recuentos más actualizados en MAPPPD —extracto hecho el 14 de septiembre de 2018, pero actualizado con datos recientes 15— correspondientes a 88 sitios existentes de reproducción de pingüinos Adelia, muestran que el 98,5 % de la población regional se ha estudiado desde 2004. Desde 2004, se ha relevado solo el 10,2 % de la población de pingüino barbijo, que en la actualidad cuenta con 233 sitios de reproducción: la mayor parte de la población de esta especie no se ha calculado en más de treinta años. En el caso del pingüino papúa, desde 2004 se ha relevado el 83,4 % de la población a través de 101 sitios de reproducción, aunque no se han hecho recuentos de la mayoría de la población restante en más de treinta años.
En la Subárea 48.2, se ha relevado el 81,4 % de la población del pingüino Adelia a través de 20 sitios desde 2004. Con respecto al pingüino barbijo, desde 2004 se ha estudiado el 46,7 % de su población a través de 79 sitios, pero no se han hecho recuentos de la mayoría de la población de esta especie en más de treinta años. Solo se ha estudiado una colonia de pingüino papúa desde 2004, que representa el 23,4 % de la población, a través de seis sitios de reproducción.
Los datos actualizados son vitales en materia de administración, incluida la gestión de pesquerías de kril, la comprensión de los impactos regionales del cambio climático y la comprensión de cómo cambian los ecosistemas, dada la recuperación en curso de los mamíferos marinos. En la actualidad, es imprescindible contar con nuevas herramientas (por ejemplo, 16, 14) para que los administradores comprendan adecuadamente las trayectorias de las poblaciones de pingüinos.
Impulsores del cambio demográfico
Se han sugerido varias hipótesis para explicar por qué cambian las poblaciones de pingüinos, entre ellas: (i) cambios del ecosistema asociados con el cambio climático regional; (ii) cambios demográficos dada la recuperación de los mamíferos marinos de la captura histórica; (iii) la pesca comercial de kril de la Antártida [Euphausia superba] y (iv) los impactos del turismo.
La variabilidad y el cambio ambientales afectan los procesos de las poblaciones de pingüinos ya sea en forma directa mediante el impacto fisiológico o en forma indirecta mediante la distribución o disponibilidad de su presa de nivel trófico medio. En escalas temporales cortas —de días a años—, el comportamiento reproductivo, así como el éxito reproductivo y las tasas de supervivencia, pueden verse afectados por eventos aleatorios —como eventos de tormentas inusuales—, la variabilidad estacional o la variabilidad interanual en las condiciones ambientales. En períodos más prolongados —de años a décadas—, los cambios persistentes pueden impulsar cambios en la distribución y abundancia, que son fácilmente detectados por programas de estudios a largo plazo (2, 17, 18).
Los efectos persistentes de la caza histórica tanto de focas como de ballenas y la pesca hacen difícil determinar la influencia del cambio climático en sí mismo (19-20). De hecho, es muy probable que no se encuentre una solución unificadora para explicar todos los cambios observados en las poblaciones de pingüinos. Es posible que diferentes impulsores del cambio afecten diversas especies y operen a diferentes escalas temporales y espaciales que van desde una sola colonia hasta la escala regional o, incluso, la escala circumpolar. Sin embargo, existe cierta evidencia convincente de que el cambio climático puede tener un efecto directo en la abundancia y en la distribución de los pingüinos. El aumento de nevadas derivado del incremento de las temperaturas y de la humedad ha favorecido las disminuciones de la población de pingüino Adelia cerca de la Base Palmer (21), fenómeno que también puede afectar a las poblaciones de las islas Shetland del Sur. Las colonias que han experimentado una mayor acumulación de nieve han disminuido más rápidamente que las colonias de pingüinos donde la erosión eólica reduce la acumulación de nieve. Del mismo modo, el pingüino papúa se reproduce en zonas con <50 % de concentración de hielo marino en la primavera austral y, tras la disminución en el hielo marino de primavera en la WAP, esta especie amplió su rango de reproducción hacia el sur en nuevas zonas (2).
Predicción y seguimiento del cambio futuro
Desentrañar los efectos del cambio climático es complejo ya que podría afectar a los pingüinos en cualquiera o en todos sus sitios de reproducción, sitios de alimentación, sitios de muda o sitios de hibernación. Es necesaria una comprensión más amplia del recorrido vital de los pingüinos y sus interacciones ecológicas. Los estudios a largo plazo de poblaciones fijas han desempeñado un papel esencial en nuestra comprensión actual de los cambios, mientras que estudios oportunos basados en embarcaciones (por ejemplo, 4) han proporcionado datos complementarios sobre la abundancia y la distribución de los pingüinos. En la actualidad, las nuevas tecnologías —tales como la teledetección remota (14, 16), los vehículos aéreos no tripulados (UAV), los dispositivos de rastreo de animales (geolocalizadores satelitales o basados en la luz) y los sistemas automatizados de identificación individual (identificación por radiofrecuencia), al igual que los sistemas de cámaras automatizadas remotas— proporcionan importantes detalles sobre la abundancia, el ciclo de vida, la demografía y la distribución terrestre y marina —alimentación y migración— a escalas locales y regionales. Se requerirá integrar esos flujos de datos dispares y analizarlos junto a los conjuntos de datos complementarios sobre la abundancia de kril —y la disponibilidad de este último para los pingüinos que buscan alimentos—, así como las condiciones ambientales (por ejemplo, hielo marino) a fin de predecir mejor el futuro de los pingüinos en toda la península antártica. Las regiones de estudio focalizado, incluidas las zonas de referencia donde está restringida la pesca, ayudarían a reducir la complejidad de los impulsores interactivos (12).
1989
Establecimiento del Programa de Seguimiento del Ecosistema de la CCRVMA (incluidos los pingüinos)
2009
Publicación de la síntesis del SCAR, Cambio Climático en la Antártida y el Medio Ambiente
2010
La Reunión de Expertos del Tratado Antártico sobre las implicaciones del cambio climático para la gestión y la gobernanza de la Antártida, celebrada en Noruega
2015
Adopción del Programa de trabajo de respuesta al cambio climático por el Comité para la Protección del Medio Ambiente
2017
Base de datos integral de acceso público de recuentos de colonias de pingüinos, MAPPPD.