



Phil N. Trathan (1), Heather J. Lynch (2)*, William R. Fraser (3)
* heather.lynch[at]stonybrook.edu
Des données probantes mettent en lumière une modification importante du nombre et de la répartition de manchots (Adélie, à jugulaire, papous, empereurs, royaux et le gorfou doré) qui se reproduisent dans la péninsule antarctique et l’arc des Antilles australes dans les sous-zones 48.1 à 48.4 de la FAO. Dans la sous-zone 48.1, le nombre de manchots Adélie et de manchots à jugulaire a décliné dans une grande partie de la péninsule antarctique occidentale (OPA) au nord de la baie Marguerite. La population de manchots Adélie est stable ou en augmentation dans la baie Marguerite ainsi que vers le sud, et stable ou en augmentation dans la péninsule antarctique orientale. À l’inverse, les manchots papous présents dans la péninsule antarctique occidentale (48.1) et dans les îles Orcades du Sud (48.2) sont plus nombreux et élargissent leur zone de reproduction vers le sud ; ailleurs dans la région, leur population est très variable mais ne présente pas de tendance identifiable. Dans la sous-zone 48.3, la population de gorfous dorés connaît un déclin considérable alors que le nombre de manchots royaux augmente. Dans la zone 48.4, les manchots à jugulaire et Adélie sont en nombre stable. Ces conclusions soulignent l’hétérogénéité considérable qui caractérise la répartition de ces espèces ainsi que l’importance de disposer de données de recensement récentes qui étayeraient les efforts pour prédire et contrôler les tendances à venir.
Il existe 18 espèces de manchots dans le monde, dont six se reproduisent régulièrement dans la péninsule antarctique et dans la mer des Antilles australes (figure 1) : Adélie [Pygoscelis adeliae], manchot à jugulaire [P. antarctica], manchot papou [P. papua], manchot royal [Aptenodytes patagonicus], manchot empereur [A. forsteri] et gorfou doré [Eudyptes chrysolophus]. Au cours du siècle dernier, les populations de manchots présentes dans la région ont évolué (p. ex. 1-2), avec des déclins marqués pour de nombreuses colonies de gorfous dorés, de manchots Adélie et de manchots à jugulaire. Par exemple, le nombre de gorfous dorés diminue dans la sous-zone 48.3 (3), et bien que les manchots Adélie et à jugulaire soient stables ou en augmentation dans la sous-zone 48.1 dans la baie Marguerite (2, 4), ils sont en diminution dans presque toute la péninsule antarctique occidentale (OPA) et jusque dans la partie septentrionale de la baie Marguerite (2). Les manchots Adélie sont en nombre stable ou en augmentation dans la région de la mer de Weddell, ainsi que dans la sous-zone 48.4 (4). Le nombre de manchots augmente fortement dans l’OPA dans la sous-zone 48.1 et sur les îles Orcades du Sud dans la sous-zone 48.2 et leur aire de reproduction s’étend vers le sud (2). Ailleurs dans la région, les manchots papous ne connaissent pas une évolution démographique significative (4), bien que d’importantes fluctuations interannuelles du nombre de manchots papous puissent compliquer l’observation de tendances. Le nombre de manchots royaux augmente dans la sous-zone 48.3 et leur aire de répartition semble s’étendre vers le sud, en direction des îles Shetland du Sud dans la sous-zone 48.1 (5), et bien que les manchots empereurs aient désormais disparu des îles Dion dans l’OPA (6), ils continuent de se reproduire à proximité de l’île Snow Hill. Ces conclusions soulignent l’énorme hétérogénéité qui caractérise la répartition de ces espèces de manchots et l’importance de mener des études comparatives à long terme dans les zones d’observation.
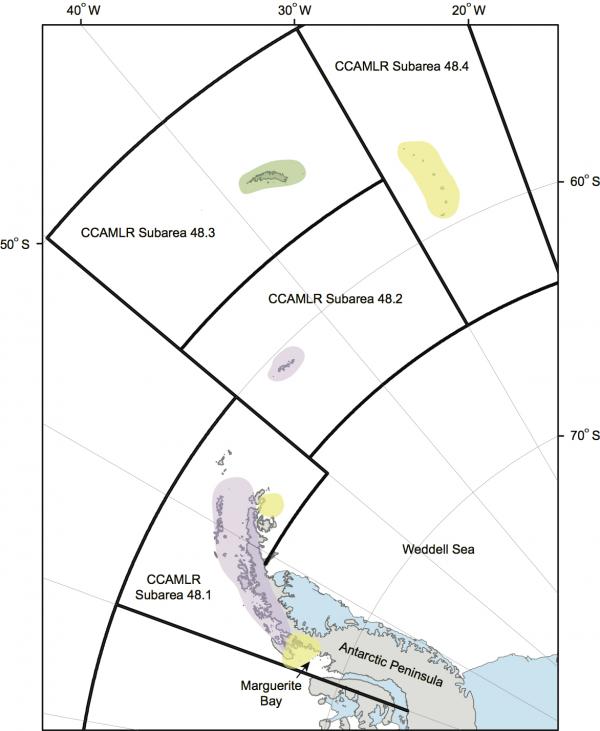
Figure 1. Péninsule antarctique et l'arc des Antilles australes. La zone en rose représente la portion de la péninsule antarctique occidentale où la population de manchots Adélie et de manchots à jugulaire décline largement et où le nombre de manchots papous augmente fortement. Les zones en jaune représentent les lieux où le nombre de manchots Adélie et de manchot à jugulaire est stable ou en augmentation (NB : aucun manchot à jugulaire n’a été observé dans la zone de la mer de Weddell ou dans la baie Marguerite). Les hachures en vert indiquent les endroits où les gorfous dorés connaissent un fort déclin. Les frontières des sous-zones statistiques établies par la FAO et utilisées par la CCAMLR sont présentées en noir.
Ensembles de données existants relatives aux manchots dans la péninsule antarctique et l’arc des Antilles australes
Les ensembles de données à long terme (20 ans ou plus) sur le nombre de manchots, leur régime alimentaire, les tendances démographiques et la phénologie proviennent notamment d’études menées sur les îles Shetland du Sud (p. ex. 1,7,8), port Lockroy (9), l’île Anvers et la pointe Biscoe (10), la pointe Cierva (11), l’île Signy (12), l’île Avian (13) et l’île Bird (3). Le programme d’inventaire des sites de l’Antarctique (4), actif dans toute la péninsule antarctique et les îles Orcades du Sud, étudie les nids et oisillons de manchots (et d’autres oiseaux de mer) depuis 1994 (4) en utilisant des navires de tourisme afin d’obtenir une couverture géographique élargie qui complète celle fournie par d’autres programmes de recherche à long terme.
Les données démographiques les plus actuelles et complètes concernant les manchots de l’Antarctique proviennent du projet MAPPPD (4). Cette base de données contient toutes les données publiées sur les populations de manchots en Antarctique, notamment des données historiques appartenant à la British Antarctic Survey, au programme d’inventaire des sites de l’Antarctique, à la division australienne de l’Antarctique, et d’autres programmes nationaux. Les données MAPPPD comprennent en outre des estimations démographiques réalisées à l’aide d’images par satellite à très haute résolution pour les manchots Adélie (14), des comptages effectués avec des drones, des photographies prises sur le terrain et des comptages en haute mer opérés à partir de bateaux.
Dans la sous-zone 48.1, les recensements les plus récents repris dans la MAPPPD (résumé réalisé le 14 septembre 2018, mais actualisé le 15 avec des données récentes) pour 88 sites de reproduction existants des manchots Adélie, montrent que 98,5 % de la population régionale a été étudiée depuis 2004. Concernant les manchots à jugulaire, parmi les 233 sites de reproduction existants, seuls 10,2 % de la population a été étudiée depuis 2004 et la plupart de la population n’a pas été comptabilisée depuis plus de 30 ans. Pour les manchots papous, 83,4 % de la population répartie sur 101 sites de reproduction ont été étudiés depuis 2004, mais pour la plupart des sites restants le recensement date de plus de 30 ans.
Dans la sous-zone 48.2, 81,4 % de la population de manchots Adélie dans 20 sites sont étudiés depuis 2004. Pour les manchots papous, 46,7 % de la population répartie sur 79 sites de reproduction ont été étudiés depuis 2004, mais la majorité de cette espèce n’a pas été étudé depuis plus de 30 ans. Des six sites de reproduction de manchots papous, seule une colonie, qui représente 23,4 % de la population, a été étudiée depuis 2004.
Il est vital de disposer de données actualisées afin d’opérer des choix de gestion éclairés, notamment pour la pêche du krill, pour comprendre les incidences du changement climatique sur la région ainsi que l’évolution des écosystèmes, étant donné l’actuel rétablissement des mammifères marins. Il devient urgent de disposer de nouveaux outils (p. ex. 16,14) si les gestionnaires veulent comprendre comme il se doit les trajectoires des populations de manchots.
Facteurs de l’évolution démographique
Plusieurs hypothèses se sont fait jour pour expliquer l’évolution des populations de manchots, dont les suivantes : (i) les changements de l’écosystème liés au changement climatique régional ; (ii) les modifications démographiques associées à la récupération des mammifères marins après des périodes de captures historiques ; (iii) la pêche commerciale du krill antarctique [Euphausia superba] ; et (iv) les conséquences du tourisme.
Les variations et les changements environnementaux affectent les populations de manchots, directement d’un point de vue physiologique ou indirectement, dans la répartition ou disponibilité de leurs proies de niveau trophique intermédiaire. Sur des échelles à court terme (quelques jours à des années), des évènements aléatoires (comme des tempêtes inhabituelles), la variabilité saisonnière, ou la variabilité interannuelle des conditions environnementales. peuvent affecter le comportement reproducteur ainsi que le taux d’efficacité de la reproduction et les taux de survie. Sur des périodes plus longues (quelques années à des décennies), les changements persistants peuvent entraîner des modifications dans la répartition et l’abondance des espèces, qui sont facilement détectées par des programmes de suivi à long terme (2, 17, 18).
Les effets persistants des pratiques séculaires de la pêche et de la chasse au phoque et à la baleine compliquent les efforts visant à déterminer l’influence du changement climatique en tant que tel (19-20). Il est en effet fort probable qu’il n’y ait pas qu’une seule raison à l’origine de tous les changements observés dans les populations de manchots. Plusieurs facteurs de changement peuvent avoir des incidences sur plusieurs espèces et agissent à des échelles spatiales et temporelles différentes qui touchent parfois une seule colonie, parfois la région tout entière ou même l’ensemble de la zone circumpolaire. Il existe néanmoins des preuves irréfutables que le changement climatique peut avoir une incidence directe sur l’abondance et la répartition des manchots. Les chutes de neige accrues résultant de conditions plus chaudes et plus humides peuvent avoir contribué aux déclins des populations de manchots Adélie à proximité de la station Palmer (21), une caractéristique qui pourrait également affecter les populations présentes dans les Shetlands du Sud. Les colonies qui ont connu plus de neiges accumulées se sont réduites plus rapidement que les colonies de manchots où le passage du vent amoindrit la couche neigeuse. De même, les manchots papous se reproduisent dans des zones ayant <50 % de concentration des glaces de mer lors du printemps austral et en raison de la diminution des glaces de mer au printemps le long de l’OPA, ils ont étendu leur aire de reproduction vers de nouvelles zones, plus au sud (2).
Prédire et surveiller les évolutions à venir
Il n’est pas aisé de faire la distinction entre les effets du changement climatique puisque cela pourrait avoir une incidence pour les manchots sur l’un ou plusieurs de leurs sites de reproduction, d’alimentation, de mue ou d’hibernation. Il est dès lors nécessaire de mieux comprendre la vie et le parcours des manchots ainsi que leurs interactions écologiques. Des études à long terme de populations fixes ont apporté des connaissances essentielles à notre compréhension actuelle des changements intervenus, alors que des études opportunistes menées depuis les navires (p. ex. 4) ont fourni des données complémentaires sur l’abondance et la répartition des manchots. Les nouvelles technologies, comme l’imagerie de télédétection (14,16), les véhicules aériens sans pilote (UAV), les dispositifs de suivi d’animaux (géolocalisateurs par satellite ou qui détectent l’intensité lumineuse), les systèmes automatisés d’identification d’individus (identification par radiofréquence) ainsi que les systèmes de vidéosurveillance automatisés, fournissent aujourd’hui des détails importants sur l’abondance, le cycle de vie, la démographie et la répartition terrestre et marine (alimentation et migration), tant à l’échelle locale que régionale. Il faudra intégrer ces flux de données disparates ainsi que leurs analyses et les ensembles de données sur l’abondance de krill (et sa disponibilité pour les manchots en période d’alimentation) et sur les conditions environnementales (p. ex. les glaces de mer) qui peuvent les compléter pour mieux prédire l’avenir des manchots dans la péninsule antarctique. Des études axées sur des régions précises, qui comprennent des zones de référence où la pêche est limitée, contribueraient à rendre moins complexe l’interaction entre les différents facteurs (12).
1989
Établissement du programme de surveillance des écosystèmes de la CCAMLR (y compris des manchots)
2009
Publication de la synthèse du SCAR, Antarctic Climate Change and the Environment
2010
Rencontre en Norvège des experts sur les conséquences du changement climatique pour la gestion et la gouvernance de l’Antarctique
2015
Adoption du programme de travail en réponse au changement climatique par le Comité pour la protection de l’environnement
2017
Base de données des recensements des colonies de manchots complète et accessible au public, MAPPPD.